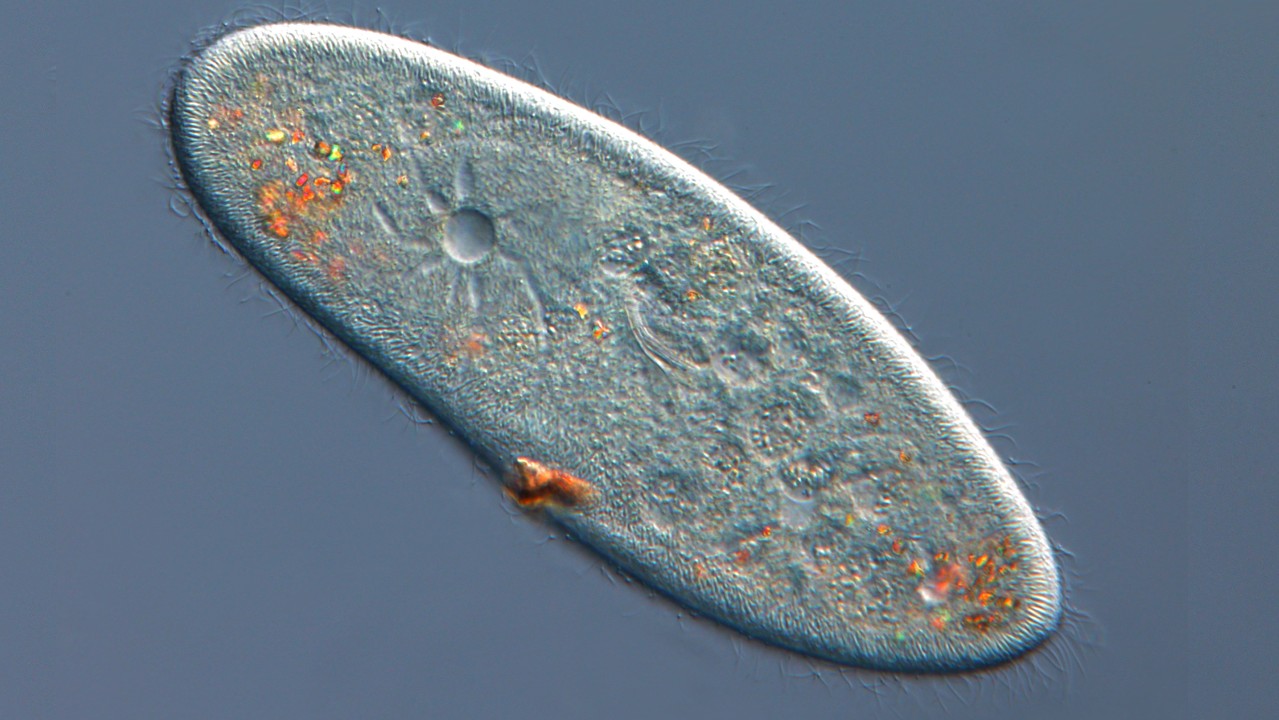
Pragecium或Paramecia是单细胞的原生物在水生栖息地中自然发现。它们通常是长方形或拖鞋形的,上面覆盖着称为纤毛的短毛结构。某些Parecia也很容易在实验室中培养,并用作有用的模型生物(一种用于了解生物学过程的非人类物种)。
外貌
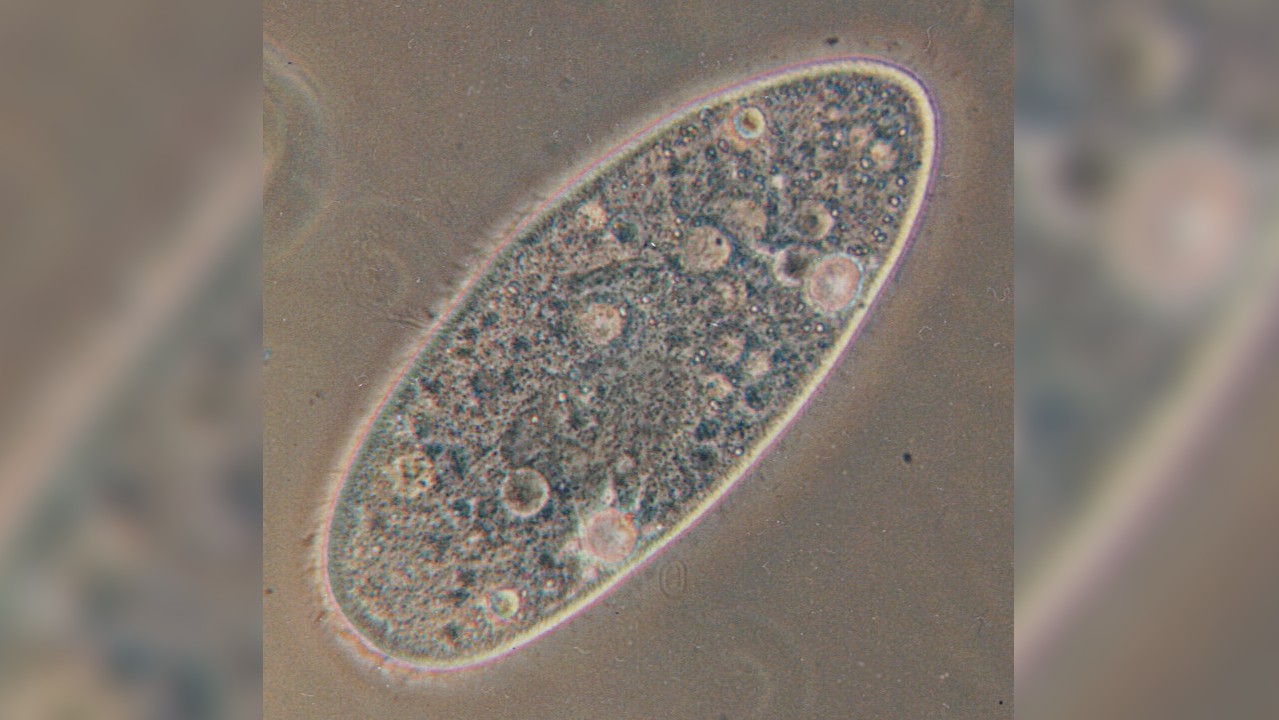
在外观上细胞伸长,并基于这种形状分为两组:Aurelia和bursaria,根据“甲虫的生物学,第二版。“(Springer,1986)。奥雷利亚的形态类型是长方形的,或“雪茄”形,后端有些锥形。另一方面,囊囊代表了“拖鞋”形状的细胞。它们往往变短,后端圆形。
依据是一组被称为纤毛的生物的一部分。加利福尼亚技术学院y。顾名思义,他们的身体被纤毛覆盖或短毛状突起。纤毛对于亚氏菌的运动至关重要。当这些结构在水生环境中来回鞭打时,它们通过其周围环境推动了生物体。
根据“根据“细胞生理来源书”(学术出版社,1995年)。有时,有机体会通过扭转纤毛击败的方向来执行“避免反应”。这导致停止,旋转或转弯,此后,黑晶石恢复向前游泳。如果多种避免反应相互跟随,则可能会使甲虫向后游泳,尽管不如向前游泳那么顺利。
西里亚(Cilia)还通过将食物推入原始的口腔开口(称为口腔凹槽)来帮助进食。 Paramecia主要以细菌为食,但已知会吃酵母,根据“甲殖的生物学”,,单细胞藻类,甚至是一些非生存的物质,例如奶粉,淀粉和粉红色的木炭。
细胞结构
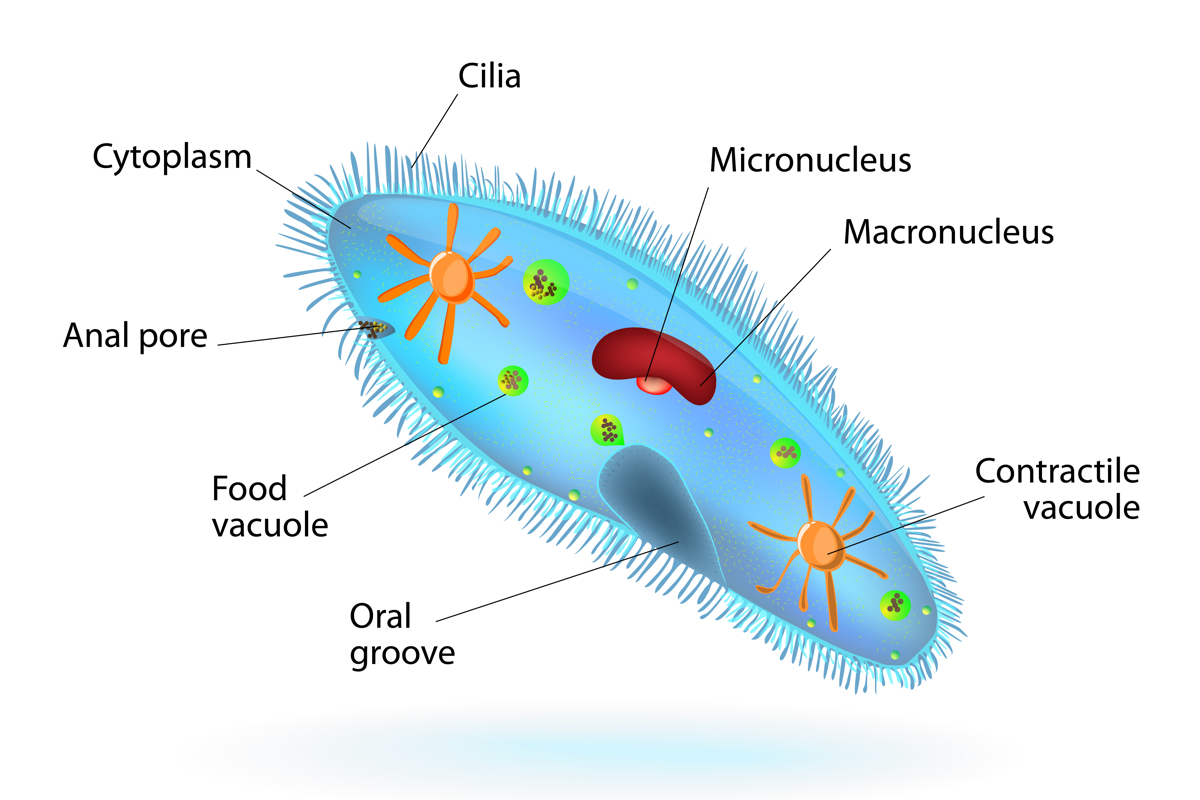
黑甲是真核生物。与原核生物相比细菌和古细菌,真核生物有组织良好的细胞。真核细胞的定义特征是存在专门的膜结合细胞机制,称为细胞器和细胞核,这是一个隔室,可容纳脱氧核糖核酸, 根据华盛顿大学。 Paramecia具有所有真核生物的许多细胞器,例如能量生成线粒体。但是,该生物还包含一些独特的细胞器。
在一个称为颗粒的外部覆盖物下,是一层有些牢固的细胞质,称为骨质。该区域由纺锤形细胞器组成,称为毛囊肿。当他们排放内容时,它们会根据“甲状腺素的生物学”而变得长,稀薄和尖刺。
毛囊肿是一种伸缩细胞器网络,用于驱除和屏蔽任何掠夺性攻击,并朝着无法预测的方向推动甲状腺素。细胞生物学杂志。杂志上发表的研究动物科学发现三尿尿素对测试的三个捕食者中的两个有效:头甲虫旋转器和Eucypris节肢动物的种类。
在胞质下方是一种更流畅的细胞质:内质。该区域包含大多数细胞成分和细胞器,包括液泡。这些是膜封闭的口袋,在动植物中,它们在植物和动物中处理废物,存储水并为细胞提供结构支持。国家人类基因组研究所液泡具有特定的功能,并具有甲苯克细胞的特定功能。食物液泡封装了甲米含量的食物。芝加哥大学。然后,他们与称为溶酶体的细胞器融合,其酶分解食物分子并进行消化形式。根据作者,收缩液泡负责渗透调节或从细胞中排出多余的水。高级生物学,第一版。“(尼尔森,2000年)。根据物种的不同,水会通过运河或较小的携带水的液泡送入收缩液泡中。当收缩液泡倒塌时,这种多余的水通过颗粒中的孔离开了甲状腺体。
也许最不寻常的特征是它们的核。 “甲虫除其他纤毛外,还具有这个相当独特的功能。”詹姆斯·福尼,普渡大学生物化学教授。 “它们有两种类型的核,它们的形状有所不同,含量和功能。”
两种类型的核是微核和大核。微生物学百科全书。微核是二倍体;也就是说,它包含每个黑质染色体的两个副本。福尼指出,小核包含有机体中存在的所有DNA。他说:“在性繁殖过程中,这是从一代人传递到另一代的DNA。”另一方面,大核包含来自微核的DNA子集。他补充说:“这是转录活性核。” “因此,是从这些mRNA中产生mRNA和蛋白质的核。”根据微生物学百科全书,大核是多倍体,或包含每个染色体的多个副本,有时甚至多达1000份。
根据福尼的说法,所有黑晶物种都有一个大核。但是,微核的数量可能因物种而异。他举例说明甲虫象物种复合物,有两个微核和黑核核核酸杆菌,有几个。
为什么存在两个不同的核?一进化原因是,这是一种机制,通过这种机制,黑晶石和其他纤毛可以避免遗传入侵者:将自己嵌入基因组中的DNA部分。福尼解释说:“就纤毛而言,有一种机制,如果在微核中有一块DNA,但它不在大核中,它将从制作的下一个大核中将其删除。” “换句话说,如果外来的某些东西进入了微核基因组,那么当制作下一个大核的情况下,它将去除,而不会包含在基因组的表达版本中。”福尼指出,某些人将其描述为原始的DNA免疫系统。也就是说,调查基因组并试图阻止入侵元素。
生殖
Paramecia可以根据其环境条件无性恋或性地再现。当有足够的营养物质可用时,在饥饿的条件下进行有性繁殖时会发生无性繁殖。此外,根据“长期饥饿的条件”,Paramecia还可以接受“自动型”或自我受精。建立地球上最复杂的结构”(Elsevier,2013年)
无性繁殖(二进制裂变)
在二进制裂变期间,一个黑晶细胞分为两个遗传上相同的后代或子细胞。根据福尼的说法,微核经历了有丝分裂,但是大核划分另一种划分,称为症状或非丝裂机制。他说:“它不是基于有丝分裂的,而是[大核]在两个细胞之间划分,并且以某种方式能够保持每个基因的副本大致相同。”
有性繁殖(共轭)
Paramecia之间的结合类似于交配。福尼说,帕拉西亚有两种交配类型,被称为奇怪甚至。这反映了以下事实:各种甲状管菌物种的交配类型用奇数甚至数字表示。例如,根据福尼的说法三尿尿素他说:“奇数与均匀的交配类型交配,但如果您是相同的交配类型,则无法交配。”此外,只有一个单元甲虫物种可以彼此交配。
在实验室条件下,该过程很容易区分。福尼说:“这些细胞结合在一起。当它们最初混合时,它们实际上可以形成相当引人注目的细胞团。” “然后那些逐渐在文化中成对的人。”
(Parmecium结合。AntonioGuillén,CC由3.0)
在有性繁殖期间,每个黑晶的微核经历减数分裂,最终将遗传含量减半以创建单倍体核。这些是在两个连接的伴侣之间交换的。来自每个伴侣融合的单倍体核,形成一个新的,遗传上变化的微核。反过来,新的微核复制以产生新的大核。
自动(自我提取)
福尼说:“自动木材本质上与共轭相同,但只有一个单元格。”在此过程中,微核重复多次复制。这些新的微核之一会重新排列其遗传含量。根据该杂志细胞和分子生命科学。
分类
一般术语“ pragecium”是指该属中的单个生物甲虫。属,是指具有相似特征的一组密切相关的生物。英国百科全书。属甲虫进一步分为称为子属的组,每个组包含一个或多个物种。
多年来,对Paramecia进行分类的方法发生了变化。最早的方法是通过视觉观察,并基于形态学,最终将所有Paramecia描述为Aurelia或Bursaria。最近,分类将形态学观察与分子和遗传信息结合在一起。根据伯克利大学。
从形态到分子系统发育学的这种转变影响了对关系中关系的理解甲虫属和物种多样性,根据MichaelaStrüder-Kypke,在加拿大安大略省圭尔夫大学分子和细胞成像设施的高级光学显微镜经理。总共有五个亚属。叶绿体,,,,直升机,,,,塞浦路斯托姆,,,,Viridoparamecium和甲虫根据期刊多样性。
Strüder-Kypke说,一种识别称为“ DNA条形码”的物种的方法已被用于甲虫。她解释说:“基于特定DNA片段的序列对物种的识别已称为DNA条形码。” “就像商店中的条形码一样,可以识别每种产品,一个足够不同的短DNA序列可以识别每个物种。”一个这样的条形码,COX1基因已被广泛用于该属甲虫,”Strüder-Kypke说。
目前有19个公认的形态学甲虫根据Strüder-Kypke的说法。她解释说,形态疗法是一种仅由独特的形态特征定义的物种,而不是遗传学或产生肥沃后代的能力。其中,15种同级物种形成了所谓的甲虫象物种综合体。根据Strüder-Kypke的说法,兄弟姐妹物种看起来相同,没有形态学的特征,但是它们在生化和遗传方面有所不同,并且无法彼此共轭。这甲虫象复杂算作单一形态学。
即使在今天,对黑晶分类法和新物种的存在的新见解仍在描述。第19个形成式,Pargecium buetschlii,在挪威的一个淡水池中发现,并在《杂志》上发表的研究中进行了描述。生物多样性与进化。同一篇论文还描述了在德国,匈牙利和巴西发现的三种新的“隐秘物种”。作者解释说,它们被视为神秘物种,因为它们很难将形态学与帕拉米氏菌的其他成员区分开。但是,其DNA [DNA条形码]中的分类标记表明它们是单独的物种。
“想法是,如果我们看着这个世界的异常栖息地或“在采样”的地区,我们仍然可能会发现新物种,”Strüder-Kypke告诉LiveCience。
用途
微生物(例如甲状腺菌)可以是监测水质的有用工具,因为来自新加坡技术大学(SUTD)发现了。为了测量海洋环境中的污染物,研究人员追踪了甲虫的运动。 “我们之所以选择Paramecia,是因为它们在水体中无处不在,并且足够大,可以用普通相机看到。”陈述。通过跟踪游泳速度和运动,研究人员能够准确检测样品中水污染物(例如重金属浓度)的质量和存在。当存在污染物(例如重金属氯化锌和硫酸铜)以及一些抗生素(例如红霉素)时,黑甲游泳速度下降。
Fernandez说:“因此,取水并测量了Paramecia的速度,可以用作一种直接的方法来评估水的可饮用性,而无需专门的设备或化学药品。” “通常,您需要为每种污染物进行不同的测试,但是Paramecia游泳是全球测量。”
其他资源
有关Pramecium的更多信息,请查看“黑质中的内共生体(微生物学专着书12)” Masahiro Fujishima和“微生物生物学”。要观看行动中的甲虫视频由YouTube频道到MicrocoSmos的旅程。
参考书目
Iwona,Rzeszutek等人,“纤毛中的编程基因组重排”,细胞和分子生命科学,第77卷,2020年5月,https://doi.org/10.1007/s00018-020-03555-2
Anne-Marie Tassin等人,“三武三尿的基础结构”,CILI,第5卷,2016年2月,2016年,https://doi.org/10.1186/s13630-016-0026-4
Van Houten J。,“ Parmecium Biology”,Evo-Devo:细胞和发育生物学中的非模型物种,第68卷,2019年10月,https://doi.org/10.1007/978-3-030-23459-1_13
Plattner H.,“黑晶中的分泌机制”,分泌机制。神经内分泌学的大师班,第8卷,2020年4月,https://doi.org/10.1007/978-3-030-22989-4_13
埃德娜(Edna) Kaneshiro,“ Amoeboid运动,Cilia和Flagella”,细胞生理学来源书,学术出版社,1995年,https://doi.org/10.1016/b978-0-12-656970-4.50051-8